
Tegenwoordig is de belangstelling voor zeldzame ziektes groter dan ooit tevoren. Er wordt meer onderzoek gedaan naar de moleculaire achtergrond en mechanismen binnen en buiten de lichaamscellen, die bij een bepaalde aandoening verstoord zijn geraakt. Door geavanceerde moleculaire technieken en het wereldwijd delen van specialistische kennis en klinische gegevens, is inmiddels een immense verzameling van genetische afwijkingen opgehelderd, welke verband houden met ziektes bij de mens. Moleculaire kennis kan bijdragen aan het verder classificeren en diagnosticeren van die ziektes. Ook zeldzame aandoeningen krijgen hierbij meer aandacht.
Een beter begrip van de zeldzame aandoening CMTC (Cutis Marmorata Telangiectatica Congenita), door het vergelijken van DNA-sequenties (het genetisch materiaal dat we bij ons dragen) van patiënten onderling, lijkt binnen handbereik. Maar zelfs als er essentiële mutaties zijn gevonden in het DNA, moet er nog een complexe puzzel worden opgelost, voordat er betekenis gegeven kan worden aan wat de gevolgen hiervan zijn voor de patiënt. Pas dan kunnen betrokken patiënten en familieleden er baat bij hebben.
Wat genetica in het algemeen betreft: DNA-moleculen, opgebouwd uit nucleotiden, vormen de bouwstenen van chromosomen en fungeren in elke lichaamscel als de basis voor vitale processen. Sommige daarvan zijn in beginsel onontbeerlijk, zoals celgroei en -ontwikkeling. Eén van de primaire functies van DNA in deze is de aanmaak van eiwitten, ook wel proteïnen genoemd. Dit zijn belangrijke bestanddelen die deel uitmaken van elke cel. Zie het als een lopende band in een fabriek, die een cel, als een levend systeem in stand houdt.
Doel van dit artikel is om duidelijk te maken wat recente ontwikkelingen in genetisch onderzoek naar afwijkingen aan het licht hebben gebracht qua verstoorde cel functies. Het wetenschappelijke artikel van Jordan uit 2019 (ref. 1) is als uitgangspunt genomen, omdat hierin corresponderende DNA-mutaties in de chromosomen van patiënten worden belicht. Patiënten die deel uitmaakten van dit onderzoek van Jordan vertoonden huidafwijkingen in combinatie met verschijnselen van onder- of overgroei, zoals ook bij CMTC-patiënten wordt gezien. Een aparte samenvatting van dit artikel van Jordan is elders op de CMTC-website (ref. 2) te vinden.
Heel in het kort, het sleuteleiwit, dat in alle geanalyseerde cases uit dit onderzoek gemuteerd bleek te zijn, is het Gα-eiwit. Dit eiwit maakt bijv. deel uit van een trimeer GPCR (een type van receptoren, dat aan een dergelijk G-eiwit gekoppeld is), waarover later meer. In dit kader is de rol van de overige twee subunits (β en γ) buiten beschouwing gelaten.
In deze rubriek komen verder, naast de genetische eigenschappen van patiënten (ofwel hun genotype), ook enkele signaleringscascades aan de orde, die in de cel signalen kunnen doorgeven, die bij de receptor zijn binnengekomen. Met als doel om een (re)actie in gang te kunnen zetten in de vorm van een cellulair proces. Mits de cascade of kettingreactie niet verstoord is geraakt door bovengenoemde DNA-mutaties en geen signalen meer kunnen worden doorgeven (‘loss of function’) óf juist teveel signaal wordt afgegeven (‘gain’ of ‘function’). Vergelijk het met onze zintuigen en het zenuwnetwerk dat daaraan gekoppeld is. Ook dit heeft als doel o.a. om ons in elke omgeving te kunnen redden middels een adequate signalering en/of (re)actie.
CMTC is geen eenduidig omschreven syndroom en kan overlap vertonen met andere, gerelateerde syndromen, zoals het Sturge-Weber-syndroom of het Adams-Oliver-syndroom (ref. 3). Een andere aandoening die overlappende symptomen vertoont met CMTC is Cutis Marmorata (CM), een breder voorkomend fenomeen dat daarentegen niet diagnostisch is (ref. 3).
De diversiteit van CMTC, als huidgerelateerd syndroom, wordt duidelijk wanneer een Google-zoekopdracht wordt uitgevoerd om te achterhalen met welke verschijnselen een CMTC-patiënt te maken krijgt. Als beschrijving van de verschijnselen bij CMTC komt voor ‘gemarmerde huid’ of ‘afgebakende plekken met gevlekte huid’ of ‘verkleurde plekken op de huid veroorzaakt door verwijde, oppervlakkig gelegen bloedvaten’ of ‘reticulair vasculair huidpatroon’ (ofwel met betrekking tot de bloedvaten netwerkvormig patroon in de huid) of ‘gespreid voorkomende huidlaesies’ enzovoort. Bovendien zijn afwijkingen aan het bewegingsapparaat, met als gevolg lichaamsasymmetrie, met soms nog aanvullende verschijnselen, kenmerkend voor dit zeldzame syndroom (ref. 4).
Sinds enkele jaren kan een subcategorie van macrocefalie-capillaire malformatie (M-CM) worden onderscheiden van CMTC als een apart syndroom, met nog steeds de suggestie, dat het een gevolg is van genetisch mozaïcisme (ref. 3). Bij een mozaïcisme wordt slechts een deel van de cellen, die tot een bepaald weefsel behoren – zoals de huid -, aangetast en het andere deel niet.
Simpelweg zou CMTC in de context van genetisch mozaïcisme als volgt kunnen worden uitgelegd (zie het kader).
CMTC is een sporadisch voorkomende (ofwel zeldzame), aangeboren aandoening. Er wordt dan ook aangenomen dat tijdens de vroeg-embryonale ontwikkeling, dus pas na bevruchting (ofwel ‘postzygotisch’), een mutatie is opgetreden in het genoom. Een dergelijke mutatie wordt een ‘de novo’ mutatie genoemd, omdat deze niet is overgeërfd, maar zich slechts in een deel van de embryonale cellen voordoet.
Dit kan onopgemerkt blijven, maar het kan ook tot uiting komen in veranderde cellulaire functies, met alle mogelijke gevolgen van dien. Als bijvoorbeeld eiwitten gevormd worden vanuit het gemuteerde gen, is het aannemelijk, dat deze eiwitten niet of minder goed zullen functioneren.
Dat slechts een deel van de cellen de mutatie blijkt te bevatten, kan komen doordat vanaf het moment van mutatie, slechts de ‘nakomelingen’ van de aangedane cel (ook wel genoemd de ‘dochtercellen’) de afwijking dragen en doorgeven.
In de groeifase worden namelijk nieuwe cellen gegenereerd door herhaalde celdeling. Daarbij worden chromosomen gekopieerd en overgenomen in de dochtercellen, inclusief dus de DNA-mutatie. Elke navolgende dochtercel die hieruit voortkomt zal de mutatie bevatten in het gekopieerde DNA, terwijl nakomelingen van de niet-aangedane cellen gevrijwaard blijven van deze mutatie.
Zo kan het mozaïekpatroon ontstaan dat in de huid van een pasgeborene met CMTC zichtbaar is (terwijl het feitelijk de onderliggende vaatafwijking (ref. 2) is die je waarneemt). Ook andere defecten, zoals lichaamsasymmetrie, kunnen het gevolg zijn van een dergelijke DNA-mutatie. Omgekeerd kan het niet uitgesloten worden, dat bij deze aandoening meerdere DNA-mutaties betrokken zijn.
Kortom, de manier waarop CMTC tot uiting komt is complex te noemen, alleen al doordat verschillende weefsels kunnen zijn aangetast bij patiënten. Daar komt bij dat de diversiteit aan vaatafwijkingen bij patiënten in de klinische praktijk, hebben geleid tot verschillende classificatiesystemen. Twee hiervan zijn door Jordan naast elkaar gezet voor de onderzochte patiëntenpopulatie (ref. 1), hetgeen tot verschillende mogelijke indelingen heeft geleid. Des te belangrijker is de rol van een patiëntenvereniging zoals CMTC/OVM (ref. 2) om ervaringen te bundelen en patiënten snel van informatie te voorzien richting diagnose en verder onderzoek.
De variatie aan omschrijvingen van het syndroom CMTC kan hiermee verklaard worden. Herschikking van gerelateerde zeldzame aandoeningen in de afgelopen jaren is tevens van invloed geweest op de naamgeving van de patiëntenvereniging. Zo maakte voorheen M-CM hier deel van uit en is de huidige benaming uitgebreid tot CMTC-OVM (overige vasculaire malformaties).
Aanscherping van het fenotypische, klinische perspectief met een verbeterd inzicht in de onderliggende moleculaire karakteristieken van aandoeningen zoals CMTC, incl. afwijkende genotypen, kan ondersteunend zijn bij diagnostisering van patiënten en wellicht aanvullende behandelmogelijkheden in de toekomst.
In het afgelopen decennium hebben onderzoeksmethoden als ‘next generation sequencing’ (NGS) DNA-mutaties aan het licht gebracht die kunnen worden aangetoond in aangetast weefsel van patiënten met gerelateerde zeldzame huidaandoeningen. Geen van de mutaties zijn vooralsnog geassocieerd met CMTC.
Een recente studie van Jordan et al. in 2019 was gericht op cases met ‘niet-gediagnosticeerde huidafwijkingen’. Twaalf van de 32 onderzochte patiënten met capillaire malformaties, vertoonden een Cutis Marmorata (CM)-achtige huidafwijking.
De onderzoekers bestudeerden twee genen, nl. het Gα11- en het GαQ-gen. Deze genen coderen voor een ander subtype van het G-proteïne α. Ze hebben aangetoond dat de cases met een CM-type significant geassocieerd waren met een mutatie in het Gα11-gen van de betreffende patiënten.
Tevens was ipsilaterale hypotrofie bij deze patiënten geassocieerd met een Gα11-mutatie. Bovendien betrof de mutatie in op één na alle gevallen het aminozuur arginine (Arg) op positie 183 van de eiwitsequentie, zowel in Gα11 (Arg183Cys of Arg183His) als in GαQ (Arg183Gln; Tabel 1).
De aanpak van deze studie werd ‘omgekeerde genotypering’ genoemd, wat duidt op een poging om genotypische kenmerken toe te kennen aan fenotypische verschijnselen bij de onderzochte patiënten. In elk van de cases werd weefsel dat niet was aangetast gebruikt als interne controle.
De bevindingen van Jordan in het kort:
| Aminozuren veranderd door mutatie | Aantal cases | G eiwit |
|---|---|---|
| Arg183Cys | 7 | GNA11 |
| Arg183His | 1 | GNA11 |
| Gln209His | 1 | GNA11 |
| Arg183Gln | 3 | GNAq |
Cutis Marmorata wordt beschouwd als een symptoom in plaats van een syndroom (ref. 3).
Gemarmerde huid is ook het meest opvallende kenmerk van CMTC. Telangiëctasieën komt bij slechts een minderheid van de patiënten voor. Dit versterkt de hypothese dat ook bij CMTC het Gα11 gen een rol kan spelen. Greene et al. suggereerde al, dat veranderingen in de MEK-cascade een rol kunnen spelen bij capillaire misvormingen (incl. GPCR, de aan het G-eiwit gekoppelde receptor). Tot nu toe wordt slechts het ARL6IP6 gen gerelateerd aan specifieke CMTC-cases (ref. 4).
Nader onderzoek zal moeten uitwijzen, of Gα11 en/of andere mutaties voorkomen in CMTC-laesies.
Eerder is een associatie met een GαQ-mutatie beschreven voor het Sturge-Weber-syndroom. Het Adams-Oliver-syndroom wordt weer in verband gebracht met mutaties in ARHGAP31 (een monomeer type G-eiwit), alsook in 5 aanvullende genen, die bij dit syndroom betrokken zijn (ref. 4,6).
Het gerelateerde Klippel-Trenauney-syndroom wordt geassocieerd met mutaties in het PIK3CA-spectrum. Van mutaties in het Gα11/Q-spectrum van G-eiwitten daarentegen, is bekend dat het tot meer diffuse syndromen leidt (ref. 5). Desalniettemin worden ook capillaire misvormingen zoals bij CMTC geschaard onder één paraplu, met de “G-eiwit receptor” mutaties, genaamd “GNA-vasculaire anomalie” (GNAVAS). En de nauw verwante Gα11/Q/14-genen staan op dit moment te boek als de belangrijkste betrokken genen (ref. 5).
Vooraleerst een korte toelichting op het fenomeen ‘signaleringscascades’. Wat hieronder verstaan wordt, is hier toegelicht aan de hand van de volgende vijf aspecten:
1. Signalering gaat middels een kettingreactie, ofwel een cascade
Belangrijk om te weten is, dat signalering via een moleculaire cascade vaak de manier is, waarop signalen in de cel kunnen worden doorgegeven. Het kan worden gezien als een vorm van communicatie, te vergelijken met het welbekende telefoonspelletje, waarbij een woord of zin wordt doorgegeven door een groep van mensen op rij. Het signaal kan onderweg verstoord raken als één van de deelnemers het signaal verkeerd doorgeeft of als het vervormd wordt. In de cel zijn het de zogeheten signaal- of boodschapper-moleculen die achtereenvolgend geactiveerd worden, ook wel te vergelijken met het doorgeven van het stokje bij een estafette.
2. Een boodschapper-molecuul werkt alleen als het juist is aangemaakt. Bij een eiwit-boodschapper is het bijvoorbeeld belangrijk dat het DNA, dat voor dit eiwit codeert, klopt
Processen die in de lichaamscellen plaatsvinden kunnen op afstand worden aangestuurd. Controle gebeurt dan van buiten de cel vandaan. Elk boodschappermolecuul onderweg naar het uiteindelijke effect is dus belangrijk, omdat ze alle deel uit maken van de ketting, als schakels die met elkaar in verbinding staan. Pas als de gehele kettingreactie in die cel goed verloopt, kan het zogeheten effectormolecuul aan het eind van de rit, zijn werk goed doen.
In de ontwikkelingsfase van het menselijk lichaam is deze aansturing belangrijk voor een juiste aanleg van de verschillende weefsels en de processen die hiervoor van belang zijn.
Aangezien veel van deze boodschappermoleculen zelf ook eiwitten zijn, is een correcte DNA-sequentie (die de code bevat voor de productie van dat eiwit) van wezenlijk belang. Net als dat voor andere eiwitten, die bijvoorbeeld als bouwsteen fungeren in de cel, belangrijk is. Onder Variatie in genotypen wordt dieper ingegaan op de DNA-code, met het Gα eiwit als voorbeeld van het boodschappermolecuul.
3. De samenwerking tussen cellen begint bij de receptor, dé ontvanger van signalen van buiten de cel
De omgeving van cellen is aan verandering onderhevig. Aan de buitenkant van de cellen zitten receptoren die bepaalde signalen kunnen oppikken. Sommige receptoren maken deel uit van de buitenlaag die om de cel heen ligt, de celmembraan. Een voorbeeld van een dergelijke receptor is de GPCR (een G-eiwit-gekoppelde receptor). Een belangrijk eiwit in de ketting van reacties die hierop volgt, is dan ook het Gα eiwit. Zodra de GPCR een signaal uit de omgeving door krijgt, wordt deze boodschap als een kettingreactie doorgegeven van molecuul tot molecuul. Het is te vergelijken met een domino, waarbij de laatste steen voor het effect zorgt. De route kan voor elk effect anders zijn. Het aantal boodschappermoleculen dat bij een route betrokken is varieert. De kans op een foutje ergens in de route, bijvoorbeeld veroorzaakt door een DNA-mutatie, is niet uit te sluiten. Steeds meer signaleringsroutes worden bekend uit wetenschappelijk onderzoek. Onderzoek naar boodschappermoleculen en mutaties daarin, is daarbij in volle gang. Zodra een mutatie bij meerdere patiënten met bepaalde verschijnselen voorkomt, kan dit klinisch relevante informatie opleveren. Om echter een werkelijk verband wetenschappelijk te kunnen aantonen, zijn veel patiënten nodig voor genetisch onderzoek.
4. Een boodschappermolecuul en het effect
Signaaltransductie gebeurt op die momenten, waarop ergens in de cel processen moeten worden uitgevoerd. Vaak betreft het signalering van buiten de cel naar de celkern waar het chromosomale DNA wordt bewaard, dat onder andere zorgt voor de eiwit-productie in de cel. Eiwitten hebben elk weer een andere functie, variërend van bouwsteen van een cel tot bijvoorbeeld een boodschappermolecuul (of eiwitten anderszins).
Een heel weefsel of orgaan zal in aanleg en ontwikkeling afhankelijk zijn van het goed functioneren van de cellen waaruit het bestaat. En dus ook van het DNA in deze cellen, waarop de codes geconserveerd liggen voor de verschillende eiwitten, waaronder de signaalmoleculen zelf.
Zie het als een domino van stenen, waarbij elke steen als signaalmolecuul fungeert. En waarbij het aanzetten van de eerste steen aan het einde van de reeks effect zal sorteren. Een effector in de kern van een cel in ontwikkeling kan bijvoorbeeld de productiemachine van nieuw gebouwde eiwitten beïnvloeden. Deze eiwitten zijn nodig voor de ontwikkeling van betreffende weefsels, waar de cel deel van uitmaakt. Met alle gevolgen vandien als er iets hapert op de route er naartoe.
Door onderling samenspel van signaalmoleculen kunnen ook aangrenzende cascades tegelijk mee worden geactiveerd. Op één signaal uit de omgeving van die cel kunnen dus meerdere effecten volgen.
5. Welke route
Om de verschillende cascades naar een bepaalde effector te kunnen onderscheiden, worden ze na ontdekking vaak genoemd naar een belangrijke factor uit die cascade. Twee belangrijke routes zijn RAS/MEK en PI3K. Factoren uit deze routes komen in het laatste hoofdstuk aan bod. Het Gα eiwit is een typisch sleuteleiwit, dat beide cascades, en daarnaast nog andere, kan beïnvloeden.
Beide genoemde cascades ofwel signaleringsroutes hebben een rol in het aansturen van cel-groei en -ontwikkeling, maar ook daarna.
Inzicht in deze mechanismen en de complexiteit ervan is een goede stap in de richting van meer begrip voor en herkenning van het scala aan verschijnselen, waarmee patiënten met een zeldzame aandoening als CMTC te maken kunnen krijgen.
Een cel is voorzien van verschillende soorten receptoren, één daarvan is de membraangebonden, G-eiwit gekoppelde receptor (GPCR). Dit type receptor communiceert zowel met de binnen- als de buitenkant van de celmembraan. GPCR’s steken namelijk integraal door de celmembraan heen.
Ze worden geactiveerd zodra een ligandmolecuul aan de buitenkant van de cel hieraan bindt. In reactie daarop wordt een signaal naar de binnenkant van de receptor afgegeven, waar het vervolgens een boodschappermolecuul kan binden en activeren, in dit geval het Gα eiwit, dat het signaal doorgeeft (Figuur 1; in groen, genaamd αGTP).
Dit Gα eiwit kan na GPCR-activatie aan GTP binden, een molecuul dat de energie levert voor activatie van fosfolipase C (PLC). Hierbij worden DAG en IP3 gevormd, welke een scala aan cellulaire processen in gang kunnen zetten. Vandaar worden het ‘tweede boodschappermoleculen’ genoemd, dus na Gα.
Uiteindelijk wordt calcium-ionen (Ca2+) vrijgemaakt en worden de proteïnekinasen C en D (PKC, D) geactiveerd. Proteïnekinasen zijn enzymen, die d.m.v. fosforylering andere eiwitten (ook wel proteïnen genoemd) kunnen activeren, zoals bijv. RAF. Veel activeringsstappen in een cascade verlopen op die manier.
Allerlei fysiologische processen binnenin de cel worden zo in gang gezet, al vanaf het eerste begin, de embryonale ontwikkeling.
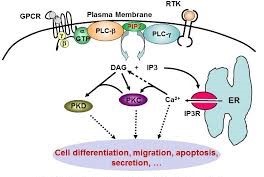
Figuur 1 – Signaleringsroute waarbij IP3 en DAG gevormd worden (ref. 7)
Naast de splitsing van PIP2 (in IP3 en DAG), kan Gα ook de PI3K-route activeren, welke aan het einde van de fosforyleringscascade resulteert in MAPK-activatie. Dit is een voorbeeld van de samenwerking tussen verschillende signaleringsroutes, die in gang gezet worden na binding van een ligand aan GPCR, als startsein (Figuur 2).
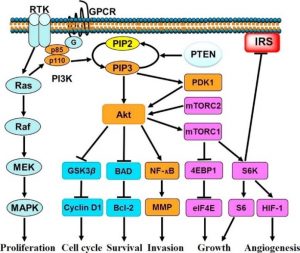
Figuur 2 – PI3K stimulatie van celgroei. PI3K, fosfoinositol-3-kinase; GPCR, G eiwit gekoppelde receptor; RTK, tyrosine kinase receptor (ref. 8).
In de studie van Jordan et al. (2019) zijn mutaties in GNA11 (een type Gα eiwit) significant geassocieerd met cases die symptomen van Cutis Marmorata vertonen.
Een vergelijkbare associatie met het Cutis Marmorata Telangiectatica Congenita (CMTC) -syndroom kan niet worden uitgesloten, alhoewel er geen aanwijzingen zijn dat dit syndroom vertegenwoordigd was in de onderzochte patiëntenpopulatie.
Auteur Dr. Marjon ten Hoor-Suykerbuyk


